全トランス型レチノイン酸に依存したLGR6の筋分化における一過的な発現は筋芽細胞の分化と融合を調節するAll-trans retinoic acid-responsive LGR6 Is transiently expressed during myogenic differentiation and is required for myoblast differentiation and fusion
論文サマリー
骨格筋を構成する筋線維は,筋形成や筋再生において分化を含む複雑な過程を経て形成される。したがって,複雑な筋形成や筋再生を理解するためには,筋分化を制御する因子を同定し,その発現機構に関する情報を得る必要がある。全トランス型レチノイン酸(ATRA)は,筋芽細胞から筋管への分化を促進する因子として機能する。我々はRNAシークエンス解析から,筋管細胞においてロイシンリッチリピート含有Gタンパク質共役受容体6 (LGR6)がATRAに応答する候補遺伝子であることを見いだした(Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118563.)。しかしLGR6の骨格筋における役割は不明である。そこでマウスC2C12筋芽細胞が筋管細胞へ分化する過程におけるLgr6 mRNAの発現パターンを評価したところ,Lgr6 mRNAの発現は分化誘導後3―6時間にかけて一過的に増加した。一方,myogenin,myomaker,myomergerのような筋分化を調節する筋原性制御因子のmRNAの発現は分化誘導後24時間で増加した。Lgr6の発現をノックダウンした細胞では分化と融合が抑制され,1つまたは2つの核を持つミオシン重鎖(MyHC)陽性細胞の割合が増加し,3つ以上の核を持つMyHC陽性細胞の割合が減少した。さらに分化中におけるLgr6 mRNAの一過的な発現上昇はATRA存在下でのみ起こった。Lgr6 mRNAの発現はATRAと同様にレチノイン酸受容体(RAR)αアゴニストとRARγアゴニストによって増加した。興味深いことに,分化誘導後3時間までLGR6を高発現させた細胞ではmyogenin,myomaker,myomergerのmRNAレベルが増加したが,分化誘導後24時間まで持続的にLGR6を高発現させた細胞ではこれらのmRNAレベルが減少した。さらに,プロテアソーム阻害剤で処理した細胞またはE3ユビキチンリガーゼのZnrf3をノックダウンした細胞では,LGR6発現がタンパク質レベルで増加した。LGR6をノックダウンした細胞では,Wnt3aの単独またはR-スポンジン2との共存によって誘導されるWnt/β-カテニンシグナル活性が減弱した。これらの結果から,LGR6の一過的な発現が筋分化促進に寄与し,その発現にはATRAが必要であることが明らかとなった(図)。さらに,タンパク質レベルでのLGR6発現は,ZNRF3に依存したユビキチン-プロテアソーム系によって負に制御され,またLGR6が筋分化中にWnt/β-カテニンシグナルを活性化すると推測された。
グラフィックサマリー
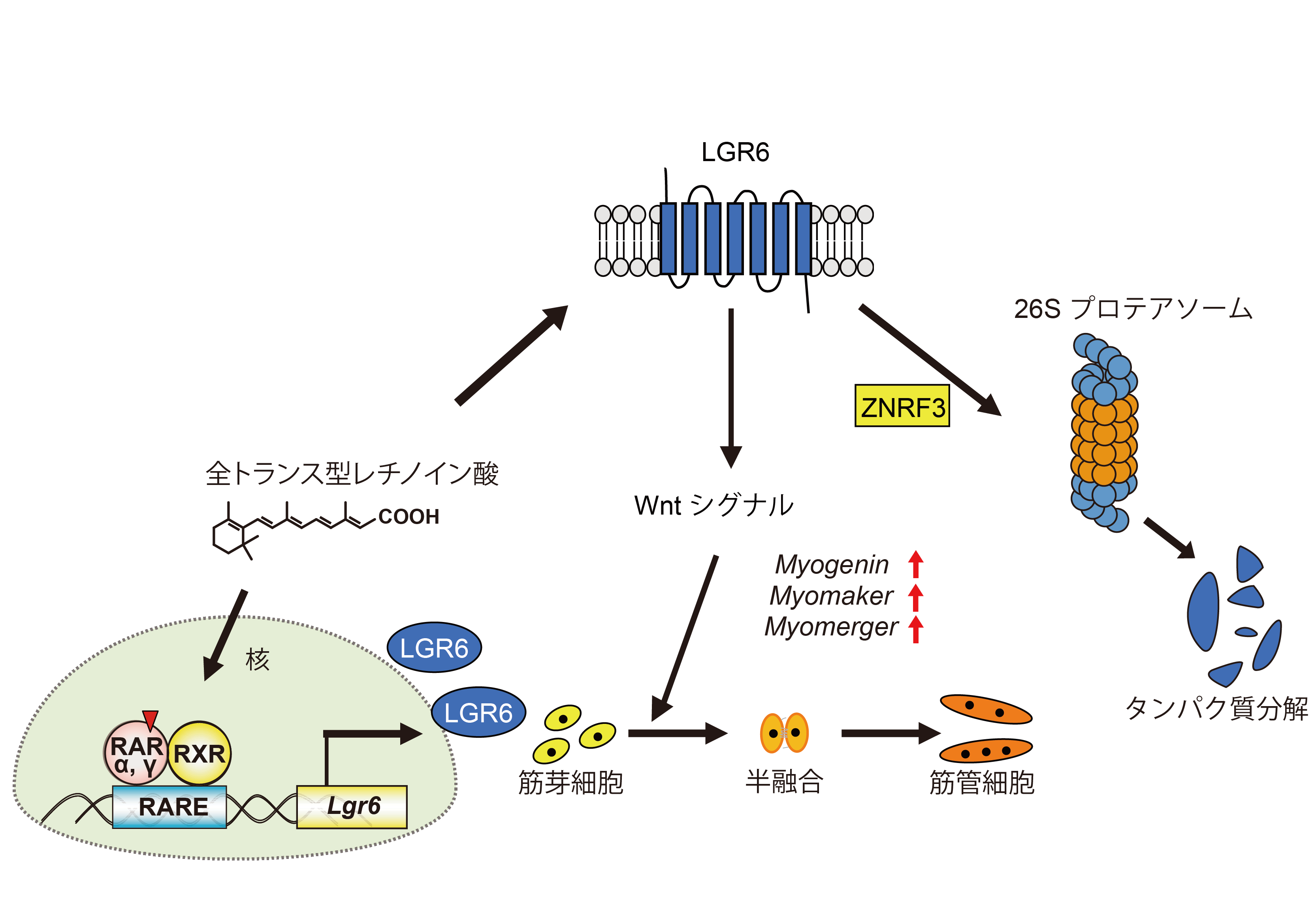
図 筋分化におけるLGR6の作用機構
解説者コメント
骨格筋量調節の一役を担っている筋分化の仕組みを理解することは、発生での筋形成や発生後での筋修復を制御できることにつながります。ビタミンAは筋分化促進因子ですが、本研究からビタミンA応答遺伝子であるLGR6が、筋芽細胞が融合する前に一過的に発現することで分化促進に寄与することが明らかになりました。今後、ビタミンAが調節する因子による筋分化制御機構をさらに詳細に検討することで、筋分化の仕組みを紐解きたいと思っています。

https://www.omu.ac.jp/agri/nc/